A Journey Through the Auditory System [สารบัญกลุ่มเรื่องที่กำลังศึกษา] เนื้อหาตอนนี้ผมสรุปจากบทที่ 4 ในชื่อเดียวกัน หนังสือ The Sense of Hearing โดย Christopher J. Plack จะเริ่มกล่าวจากหัวข้อ 4.2 The Cochlea ละหัวข้อแรก 4.1 From Air to Ear ซึ่งได้พูดถึงไปก่อนหน้านี้ในบล็อกหลาย ๆ ตอนแล้ว สำหรับเนื้อหาสรุปของบทที่ 2 และ 3 ดู The Nature of Sound (สรุป) กับ Production, Propagation, and Processing of Sound (สรุป) อนึ่ง ลำดับเนื้อหาที่จะสรุปในตอนนี้ ผมไม่เรียงเป็นข้อตามส่วน Summary เหมือน 2 บทแรก แต่เรียงตามลำดับหัวข้อของหนังสือ The Cochleaที่ cochlea ของหูชั้นในนี่แหละครับที่การสั่นของคลื่นเสียงถูกแปลงเป็นสัญญาณไฟฟ้า (electrical neural activity) cochlea เป็นท่อขนาดเล็กมีของเหลวอยู่ภายใน ยาวประมาณ 3.5 เซ็นติเมตร เส้นผ่านศูนย์กลางเฉลี่ย 2 มิลลิเมตร อันที่จริง เส้นผ่านศูนย์กลางจะค่อย ๆ เล็กลงตามระยะห่างจากหน้าต่างวงรีหรือบริเวณที่เรียกว่า base โดยเส้นผ่านศูนย์กลางเล็กสุดที่ apex ท่อนี้ขดเป็นก้นหอยประมาณ 2.5 รอบ ผนังแข็งนะครับคล้ายกระดูก ฉะนั้น เราจะจับ cochlea ที่ขดเป็นก้นหอยออกมาคลี่เป็นเส้นตรงไม่ได้ รูปด้านล่างแสดงภาพตัดขวางของ cochlea เห็นว่าท่อตามแนวยาวถูกแบ่งโดย membrane สองอัน ได้แก่ Reissner's membrane กับ basilar membrane ทำให้เกิดห้องหรือช่องว่างหรือท่อย่อยซึ่งเต็มไปด้วยของเหลว 3 ส่วน มีชื่อเรียก scala vestibuli, scala media กับ scala tympani ซึ่ง s. vestibuli กับ s. tympani เชื่อมต่อกันผ่านช่องเปิดเล็ก ๆ เรียก helicotrema ระหว่าง basilar membrane กับผนังของ cochlea ที่ apex (ดูรูป) ส่วน media เป็นท่อย่อยที่แยกออกไปต่างหาก มีส่วนประกอบของเหลว (endolymph) แตกต่างจากอีกสองท่อย่อย (perilymph)   รูปต่อมาเป็นภาพตัดขวางขยายโครงสร้างที่เป็นเจลาตินบน basilar membrane เรียกว่า tectorial membrane ซึ่งมี organ of Corti อยู่ระหว่าง membrane ทั้งสอง organ of Corti ประกอบด้วย hair cells หลายแถว (hair cells เป็นเซลล์ชนิดพิเศษชนิดหนึ่ง พวกมันจะมีส่วนคล้ายขนเล็ก ๆ เรียกว่า stereocilia ยื่นออกมา) และเซลล์สนับสนุนแบบต่าง ๆ รวมถึงปลายประสาท (nerve endings) ใน cochlea ของคน จะมี hair cells หนึ่งแถวที่อยู่ด้านใน เรียกว่า inner hair cells และด้านนอกมีได้มากถึง 5 แถว เรียก outer hair cells ตลอดความยาว cochlea คาดว่ามี hair cells ภายในราว 3,500 เซลล์ และภายนอกราว 12,000 เซลล์ และ ปลายสุดของ stereocilia ของ outer hair cells จะทิ่มฝังอยู่ใน tectorial membrane (ขณะที่ hair ของ inner ไม่ฝังนะครับ) หน้าที่ของ outer hair cells คือ เปลี่ยนสมบัติเชิงกลของ basilar membrane (รายละเอียดจะพูดถึงในบล็อกตอนอื่น) ส่วนหน้าที่ของ inner hair cells คือ แปลงการสั่นของ basilar membrane ไปเป็นสัญญาณไฟฟ้า  The Basilar Membraneเสียงเข้ามายัง cochlea ผ่านทางหน้าต่างวงรี (มันคือ membrane อันหนึ่งที่ปิดกั้นระหว่างหูชั้นกลางกับ cochlea) ของเหลวใน cochlea แทบไม่สามารถบีบอัดได้ ดังนั้นเมื่อหน้าต่างวงรีถูกผลักเข้ามาใน cochlea เนื่องจากการสั่นของกระดูกโกลน ทั้ง Reissner's membrane และ basilar membrane จะถูกผลักลง และหน้าต่างวงกลมซึ่งอยู่อีกด้านหนึ่งของ base จะถูกผลักออก นั่นคือ การสั่นของกระดูกโกลนทำให้ basilar membrane สั่นbasilar membrane เป็นส่วนสำคัญต่อการรับรู้เสียงในสัตว์เลี้ยงลูกด้วยนม มันทำหน้าที่แยกองค์ประกอบทางความถี่ของเสียง บริเวณ base ใกล้กับหน้าต่างวงรีนั้น basilar membrane แคบและแข็งทื่อ บริเวณดังกล่าวนี้อ่อนไหวต่อความถี่สูง ส่วนที่ปลายอีกด้านหนึ่ง บริเวณ apex จะกว้างกว่าและไม่แข็งทื่อเท่า จึงเป็นบริเวณที่อ่อนไหวต่อความถี่ต่ำ (ดูรูปข้างบน จะเห็นว่า basilar membrane กว้างขึ้นเมื่อ cochlea มีขนาดเล็กลง) สมบัติอันนี้ของ membrane เปลี่ยนแปลงอย่างต่อเนื่องจาก base ถึง apex ดังนั้น แต่ละตำแหน่งบน basilar membrane จะอ่อนไหวต่อความถี่เฉพาะบางค่า (เราเรียก characteristic frequency) ตัวอย่างเปรียบเทียบเพื่อให้เข้าใจกลไกการทำงานของ basilar membrane ให้นึกถึงสปริงแขวนเป็นแถวยาวเรียงกันหลายอันบนท่อนไม้ที่วางในแนวขนานกับพื้นโลก โดยสปริงแต่ละอันผูกติดอยู่กับมวลก้อนหนึ่ง สปริงที่อยู่ซ้ายมือสุดแข็งทื่อสุด (stiff) (ตอนเรียน ม.ปลาย stiffness ของสปริงเราจะแสดงด้วยค่าคงที่สปริงหรือค่านิจสปริง k ซึ่งความถี่ของการเคลื่อนที่แบบซิมเปิ้ลฮาร์มอนิกของมวลติดสปริงยกกำลังสองจะแปรตาม k และแปรผกผันกับมวล) และความแข็งทื่อของสปริงแต่ละอันที่แขวนอยู่นั้น ค่อย ๆ ลดลงเรื่อย ๆ เมื่อสปริงแขวนใกล้ปลายทางด้านขวามากขึ้น ๆ นั่นคือ สปริงขวามือสุด เป็นสปริงที่หลวมหรือคลายที่สุด อันนี้เรากำลังเปรียบเทียบให้ระบบสปริงติดมวลบนท่อนไม้เป็น basilar membrane โดยซ้ายมือคือ base และขวาคือ apex สมมติว่าเรามีสปริงอันหนึ่ง แล้วผูกปลายด้านหนึ่งของสปริงติดกับมวล แล้วปล่อยให้มันเคลื่อนที่ขึ้นลงเด้งดึ๋ง ๆ เองสักพักตามธรรมชาติก่อนหยุดนิ่ง อัตราการสั่นของมวลนั่นแหละครับคือความถี่เรโซแนนต์ของระบบ ถ้าสปริงแข็งทื่อมาก มวลก็จะเด้งดึ๋งด้วยความเร็วสูง นั่นคือ ความถี่มาก แต่ถ้าสปริงคลาย (loose) มวลก็จะเคลื่อนที่ขึ้นลงช้า มีความถี่ต่ำ ทีนี้ ถ้าคุณใช้มือจับปลายอีกข้างหนึ่งของสปริง แล้วเคลื่อนมือขึ้นลงด้วยความถี่สูงกว่าความถี่เรโซแนนต์ของระบบ มวลก็จะสั่นไม่มาก แต่ถ้ามือของคุณเคลื่อนที่ขึ้นลงด้วยความถี่เรโซแนนต์ การสั่นก็จะมาก กลับมาที่แท่งไม้ซึ่งมีสปริงติดมวลผูกเรียงเป็นแนวยาว ถ้าเราจับแท่งไม้นั้น ถือในแนวขนานกับพื้นโลก แล้วเคลื่อนแท่งไม้ขึ้นลงด้วยความถี่ค่าหนึ่ง สิ่งที่เราจะเห็นคือ มีมวลกลุ่มหนึ่งสั่นมากกว่ามวลที่เหลือ ทั้งนี้เป็นเพราะ ความถี่ของการเคลื่อนแท่งไม้ขึ้นลงใกล้เคียงกับความถี่เรโซแนนต์ของมวลกลุ่มนั้น ถ้าความถี่ของท่อนไม้มีค่าน้อย ๆ มวลที่สั้นแรงจะอยู่ทางขวา (apex) ของท่อนไม้ แต่ถ้าความถี่ของท่อนไม้มาก มวลจะอยู่ทางซ้าย (base) ทีนี้ ถ้าคุณสามารถเคลื่อนแท่งไม้ขึ้นลงในรูปแบบที่มีความถี่สองค่ารวมกัน (ทำได้นะครับ นึกภาพมือจับแท่งไม้สั่น ๆ ด้วยความถี่สูง ขณะเดียวกันแขนก็ยกมือสั่น ๆ นั้นขึ้นลงด้วยความถี่ต่ำ) การสั่นของมวลทางด้านซ้ายจะเป็นเพราะความถี่สูง ส่วนการสั่นของมวลทางขวาจะเป็นเพราะความถี่ต่ำ นั่นเสมือนกับระบบนี้ของเราสามารถแยกความถี่ที่ผสมกันอยู่ออกมาได้ นี่เป็นเพียงภาพง่าย ๆ ของกลไกแยกองค์ประกอบทางความถี่ ขณะที่ basilar membrane กับ cochlea มีความซับซ้อนมากกว่านี้เยอะ การเคลื่อนที่ของ basilar membrane ขึ้นอยู่กับความเฉื่อยของของเหลวที่อยู่รอบ ๆ, เรโซแนนซ์ใน tectorial membrane, stereocilia ของ outer hair cells และส่วนสำคัญคือ กิริยาเชิงกล (mechanical action) ของ outer hair cells (รายละเอียดส่วน outer hair cells ช่วย basilar membrane อย่างไร จะกล่าวถึงในบล็อกตอนอื่น) จากกลไกที่กล่าวมา ทำให้เราพูดได้ว่า basilar membrane ทำตัวเหมือนกับ band-pass filter หลาย ๆ อัน (bank) ที่มีช่วง bandwidth ความถี่ปล่อยผ่านซ้อนทับกัน ดังรูป 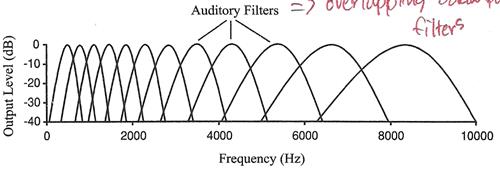 แต่ละตำแหน่งบน basilar membrane มี characteristic frequency, bandwidth และ impulse response เฉพาะตัว ตอนที่เสียงซึ่งมีความถี่รวมกันแบบซับซ้อนเดินทางเข้ามาในหู องค์ประกอบความถี่สูงของเสียงนั้นจะกระตุ้น basilar membrane แถว ๆ base ขณะที่องค์ประกอบความถี่ต่ำจะกระตุ้นแถว ๆ apex ดังนั้น basilar membrane จึงเหมือนกับตัววิเคราะห์สเปกตรัมของเสียงที่เข้ามา ดังรูป  characteristic frequency ของตำแหน่งต่าง ๆ บน basilar membrane ไม่ได้เปลี่ยนแปลงอย่างเป็นเชิงเส้นจาก base ถึง apex แต่ค่อนข้างเป็น logarithm นะครับ ความถี่สูงจะอยู่ใกล้ชิดกันมากกว่าความถี่ต่ำ ดังรูป  The Traveling Waveถึงแม้ basilar membrane จะมีขนาดเล็ก ส่วนที่กว้างที่สุดแค่ 0.45 มิลลิเมตรเท่านั้น แถมยังอยู่ในผนังแข็งคล้ายกระดูกของ cochlea แต่นักวิทยาศาสตร์ก็สามารถศึกษาการเคลื่อนที่ของ basilar membrane ในการตอบสนองต่อเสียงได้โดยตรงจากการผ่าตัด และคนแรกที่บุกเบิกเรื่องนี้คือ Georg von Békésy (ได้รับรางวัลโนเบลสาขาแพทย์ปี 1961 จากเรื่องนี้แหละครับ) เขาศึกษาการเคลื่อนที่ของ basilar membrane ใน cochlea ที่ตัดออกมาจากศพคนและสัตว์ อันที่จริงเขาดูการเคลื่อนที่ของอนุภาคเงินที่โปรยกระจายอยู่บน Reissner's membrane ซึ่งมันก็เคลื่อนที่ไปพร้อมกับโครงสร้างรอบ ๆ scala media รวมถึง basilar membrane และ organ of Corti ฉะนั้นการเคลื่อนที่ของอนุภาคเงินเหล่านี้ก็สามารถใช้บอกการเคลื่อนที่ของ basilar membrane ได้ด้วย  von Békésy สังเกตว่า ถ้าเราป้อน pure tone เข้าหู รูปแบบลักษณะเฉพาะของการสั่นจะเกิดขึ้นบน basilar membrane ถ้าเรานึกภาพคลี่ cochlea ให้เป็นท่อตรงยาว การเคลื่อนที่ของ basilar membrane จะดูคล้ายคลื่นน้ำเดินทางจาก base ไปยัง apex รูปแบบการสั่นนี้เรียกว่า traveling wave ดังรูปข้างบน ถ้าเรามองตาม traveling wave จาก base ไปยัง apex เราจะเห็นว่าคลื่นค่อย ๆ โตขึ้นกระทั่งถึงจุดสูงสุด อันเป็นจุดบน basilar membrane ที่มีเรโซแนนซ์ที่ความถี่ของ tone ก่อนคลื่นจะเล็กลงและหายไปอย่างรวดเร็ว โปรดระลึกว่า คลื่น traveling นี้ เป็นผลสืบเนื่องจากการที่แต่ละตำแหน่งบน basilar membrane เคลื่อนที่ขึ้นและลงในการตอบสนองต่อการกระตุ้นด้วย pure tone โดยที่ความถี่ของการสั่นแต่ละตำแหน่งนั้นจะเท่ากับความถี่ของ pure tone เรื่องที่เข้าใจผิดกันบ่อยเรื่องหนึ่งคือ การเคลื่อนที่ของ traveling wave จาก base ถึง apex เป็นผลจากการเปลี่ยนแปลงความดันที่เข้ามายัง cochlea ผ่านหน้าต่างวงรีที่ base อันนี้ผิดนะครับ เพราะเสียงเคลื่อนที่เร็วมากในของเหลวใน cochlea ฉะนั้น ทุกจุดบน basilar membrane จึงได้รับการกระตุ้นแทบจะพร้อม ๆ กันตอนที่มีความดันเปลี่ยนแปลงเกิดขึ้นที่หน้าต่างวงรี การเคลื่อนที่แบบลักษณะเฉพาะดังกล่าวของ traveling wave เกิดขึ้นเพราะมีการหน่วงเฟส (phase delay) ที่เพิ่มขึ้นจาก base ถึง apex ทำให้ membrane ที่ apex สั่นช้ากว่าที่ base และการหน่วงนี้เป็นผลจากลักษณะทางกายภาพของ membrane ที่ส่วน base มีความแข็งทื่อและแคบกว่าส่วน apex (นั่นคือ ระบบ stiffness-limited จะตอบสนองเร็วกว่าระบบ mass-limited) รูปต่อไปนี้แสดง snapshot ของ membrane ขณะตอบสนองต่อ pure tone สองตัว ที่ความถี่ 2 kHz และ 200 Hz ตามลำดับ เห็นว่าช่วงความกว้างการตอบสนองของความถี่ต่ำบน membrane จะกว้างกว่าความถี่สูง (ดูความยาวของ envelope) เพราะแต่ละตำแหน่งบน basilar membrane จะทำตัวเหมือน band-pass filter ที่ย่านความถี่ผ่านแตกต่างกันไป ยิ่งความถี่ต่ำ ก็เหมือนมันผ่าน filter ได้เกือบทุกตัวบนด้วยการลดทอนที่ไม่เท่ากัน เราจึงเห็นจากรูปว่า basilar membrane เกือบทั้งสายตอบสนองต่อความถี่ต่ำ ขณะที่มีบริเวณช่วงไม่กว้างนักตอบสนองต่อความถี่สูง  Transduction & How Do Inner Hair Cells Work?ภารกิจแปลงการสั่นของ basilar membrane ไปเป็นสัญญาณไฟฟ้าเพื่อส่งต่อให้สมองเป็นหน้าที่ของ inner hair cells ด้านบนของ hair cells มี stereocilia อยู่หลายแถว หน้าตาคล้ายขนเส้นเล็ก ๆ ตอนที่ basilar membrane กับ tectorial membrane เคลื่อนที่ขึ้นลงนั้น ขนพวกนี้จะถูกทำให้เอียงเฉียงด้านข้าง ดังรูป ในความเป็นจริง stereocilia เอียงแค่นิดเดียว (รูปที่เห็นถูกทำให้เว่อร์ไปนิดนะครับ) สำหรับเสียงระดับใกล้เคียงขีดเริ่มการได้ยิน พวกมันเอียงไปด้วยค่าการกระจัดแค่ 0.3 นาโนเมตรเท่านั้น พูดว่าถ้า stereocilia มีขนาดเท่ากับตึก Sears ในชิคาโก้ ระยะดังกล่าวก็เท่ากับการกระจัดของยอดแค่ 5 เซ็นติเมตร  stereocilia แต่ละเส้นจะเชื่อมโยงกันด้วยเส้นใยโปรตีน เรียกว่า tip link ตอนที่ stereocilia เอียงเข้าหา scala media หรือชี้ออกนอก cochlea เส้นใย tip link นี้จะถูกดึงให้ยืด เปิดช่องเข้าเซลล์เล็ก ๆ ดังรูป  ตอนที่ช่องนี้เปิดออกนั้น K+ (โปรตัสเซียมไอออน) จะไหลเข้าไปใน hair cell เพิ่มศักดาไฟฟ้าของเซลล์ (ในระดับ mV) และเนื่องจากศักดาไฟฟ้านิ่ง (resting electric potential) ของ inner hair cell มีค่าเป็นลบ (ประมาณ -45 mV) เราจึงเรียกการเพิ่มขึ้นของศักดาไฟฟ้าดังกล่าวว่า depolarization และกระบวนการ depolarization นี่เองที่ทำให้สารเคมีสื่อประสาท (neurotransmitter) ถูกปล่อยเข้าช่องเล็ก ๆ (synaptic cleft) ระหว่าง inner hair cell กับเซลล์ประสาทในเส้นประสาทรับรู้เสียง ดังรูป  ตอนที่สารสื่อประสาทแพร่มาถึงเซลล์ประสาท (นิวรอน) มันจะทำให้เกิดพัลซ์กระแสไฟฟ้า (spike) ขึ้นในเซลล์ประสาท ถ้า stereocilia เอียงไปในทิศตรงข้าม หรือชี้เข้าหาศูนย์กลางของ cochlea นั่นคือ tip link คลาย ไม่ถูกดึง ทำให้ช่องทางดังกล่าวปิด ลดการปลดปล่อยสารสื่อประสาท ยิ่งการเคลื่อนที่ของ basilar membrane มากเท่าไร tip link ก็ยิ่งถูกดึงให้เปิดช่องกว้างมากขึ้นเท่านั้น และยิ่งมีการเปลี่ยนแปลงทางไฟฟ้าใน hair cell มาก สารสื่อประสาทก็จะยิ่งถูกปล่อยมาก สัญญาณไฟฟ้าในเส้นประสาทมากตามด้วย สำหรับ outer hair cell ก็ถูกกระตุ้นด้วยวิธีเปิดปิดช่องผ่านไอออนเช่นเดียวกับ inner cell แต่คิดกันว่า การเปลี่ยนแปลงศักดาไฟฟ้าของเซลล์นี้จะทำให้ความยาวของเซลล์เปลี่ยน ดังนั้นมันจึงก่อให้เกิดผลประทบต่อ basilar membrane (จะกล่าวถึงในบทต่อไป) ข้อสำคัญ outer hair cell ไม่ข้องเกี่ยวกับการส่งผ่านข้อมูลไปยังสมอง The Auditory Nerveคำถาม สัตว์ขนาดใหญ่จะส่งข้อมูลจากส่วนหนึ่งไปยังอีกส่วนหนึ่งของร่างกายได้อย่างไร คำตอบ ระบบประสาท ระบบประสาทประกอบด้วยเซลล์ประสาทหรือนิวรอนซึ่งทำหน้าที่ติดต่อสื่อสารอย่างรวดเร็วระหว่างเซลล์รับสัมผัส (sensory cell) เซลล์กล้ามเนื้อ และสมอง สมองคนเรามีนิวรอนมากกว่าหนึ่งแสนล้านเซลล์ แต่ละเซลล์มีการเชื่อมต่อกับเซลล์อื่นอีกนับร้อย นิวรอนกับการเชื่อมต่อนี่แหละครับเป็นโครงข่ายประมวลผลที่ซับซ้อนและมีกำลังมาก ถึงขั้นทำให้เราคิดได้ รู้สึกได้ และดูทีวีได้4 ส่วนประกอบหลักของนิวรอนได้แก่ dendrite, soma (ลำตัวเซลล์), axon กับ terminal button ดังรูป 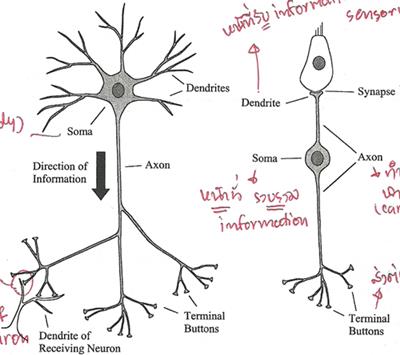 รูปนี้แสดงให้เห็นโครงสร้างของนิวรอน 2 แบบ รูปซ้ายเป็นนิวรอนที่พบเห็นได้ในสมอง มี dendrite กับ terminal button หลายเส้น ที่มุมล่างซ้ายของรูปซ้ายแสดง terminal button สร้าง synapse กับ dendrite ของนิวรอนอื่น ส่วนรูปขวาเป็นนิวรอนรับสัมผัส เช่น ตัวที่เชื่อมต่อกับ hair cell ใน cochlea พวกนี้มี dendrite เดียว พูดคร่าว ๆ หน้าที่ของ dendrite คือ รับข้อมูลจากเซลล์รับสัมผัส อย่าง inner hair cell หรือจากนิวรอนตัวอื่น, soma รวบรวมข้อมูลข่าวสาร, axon นำพาข้อมูลข่าวสาร และ terminal button ส่งต่อข้อมูลให้กับ dendrite ของนิวรอนตัวอื่น synapse คือ การเชื่อมระหว่าง terminal button กับ dendrite หรือระหว่างเซลล์รับสัมผัสกับ dendrite โดยทั่วไปในสมองของเรา dendrite ของนิวรอนตัวหนึ่งจะสร้าง synapse กับ terminal button ของนิวรอนตัวอื่นอีกหลายร้อยตัว ความยาวของ axon อาจยาวได้เกือบ 1 เมตรในนิรอนที่เกี่ยวข้องกับการเคลื่อนไหวของกล้ามเนื้อ มันนำพาข้อมูลข่าวสารในรูปอิมพัลซ์ไฟฟ้า เรียกว่า action potential หรือ spike โดยขนาดของ spike คงที่ประมาณ 10 mV แต่รูปแบบของ spike หรือจำนวนความถี่ spike ต่อหน่วยเวลา หรือที่เรียกว่า firing rate จะขึ้นอยู่กับข้อมูลข่าวสาร อัตราเร็วของ spike ใน axon อาจสูงถึง 120 m/s การเปลี่ยนแปลงศักดาไฟฟ้าที่ terminal button อันเนื่องจากมี spike เดินทางมาถึงนั้น จะทำให้มีการปลดปล่อยสารสื่อประสาท ซึ่งจะแพร่กระจายผ่านช่อง synapse ระหว่างเซลล์ ยิ่งมี spike มาถึงถี่ขึ้น ก็ยิ่งปล่อยสารสื่อประสาทมากขึ้น เมื่อนิวรอนฝั่งรับตรวจจับสารสื่อประสาทนั้นได้ มันอาจจะทำให้เกิด spike ในตัวมัน (หรืออาจจะยับยั้ง spike ที่กำลังเกิดอยู่ในตัวมันก็ได้ แล้วแต่กรณี) นั่นคือ การสื่อสารดังกล่าวเป็นกระบวนการ electrochemical Activity in the Auditory Nerveเส้นประสาทการรับรู้เสียงคือกลุ่มของ axon (หรือใยประสาท) ที่เชื่อมกับ (หรือพูดว่า สร้าง synapse กับ) hair cell ในคนเรามีประมาณ 30,000 นิวรอน และเส้นใยประสาทเหล่านี้ส่วนใหญ่จะเชื่อมกับ inner hair cell โดยที่ inner hair cell แต่ละตัวจะเชื่อมกับ dendrite ประมาณ 20 เส้นใยประสาทเนื่องจาก inner hair cell แต่ละตัวติดกับตำแหน่งเฉพาะตำแหน่งใดตำแหน่งหนึ่งบน basilar membrane ดังนั้น นิวรอนแต่ละตัวในเส้นประสาทการรับรู้เสียงจึงนำพาข้อมูลเกี่ยวกับการสั่นของ basilar membrane ที่ตำแหน่งเพียงตำแหน่งเดียวใน cochlea ทีนี้ แต่ละตำแหน่งดังกล่าวอ่อนไหว (sensitive) ต่อความถี่ลักษณะเฉพาะที่เจาะจงค่าหนึ่ง ทำให้ เราพูดได้ว่านิวรอนแต่ละตัวก็อ่อนไหวต่อความถี่ลักษณะเฉพาะบางค่านั้นเช่นกัน รูปด้านล่างแสดงให้เห็นว่านิวรอนแต่ละตัวจะมีความอ่อนไหวลดลงอย่างรวดเร็วเมื่อความถี่เสียงที่มากระตุ้นออกห่างจากความถี่ลักษณะเฉพาะ 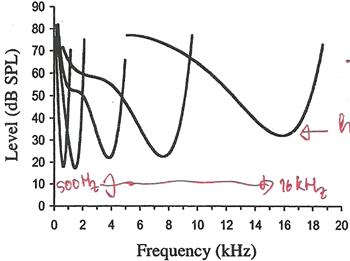 กราฟในรูปนี้แสดงความสัมพันธ์ระหว่างระดับของ pure tone ที่ต้องป้อนเข้าหูเพื่อให้ firing rate ของนิวรอน 5 ตัว (แต่ละกราฟเป็นของนิวรอนคนละตัวกัน) เพิ่มขึ้นในปริมาณที่เริ่มตรวจวัดได้ เราเรียกกราฟนี้ว่า frequency threshold tuning curve (รูปมาจากเส้นประสาทรับรู้เสียงของหนู chinchilla) จุดต่ำสุด คือ จุดที่อ่อนไหวที่สุด ถึงแม้ไม่มีเสียง เส้นใยส่วนใหญ่ก็มี firing rate ระดับพื้น เรียกว่า spontaneous activity ซึ่งราว 90% ของเส้นใย จะมี spontaneous rate สูงประมาณ 60 spike ต่อวินาที เส้นใยเหล่านี้มีแนวโน้มค่อนข้างอ่อนไหวและเพิ่ม firing rate เมื่อถูกกระตุ้นที่ระดับต่ำ เส้นใยที่เหลืออีก 10% มี spontaneous rate ราว 10 spike ต่อวินาที เป็นพวกที่มีความอ่อนไหวน้อยกว่า ความอ่อนไหวที่แตกต่างกันนี้คาดว่าอาจสัมพันธ์กับตำแหน่งของ synapse กล่าวคือ ถ้า synapse อยู่ใกล้กับ outer hair cell มันจะเป็นเส้นใยที่มี spontaneous rate สูง แต่ถ้า synapse อยู่อีกด้านหนึ่ง จะมี spontaneous rate ต่ำ พอนิวรอนถูกกระตุ้นด้วย pure tone ที่ตรงกับความถี่ลักษณะเฉพาะของมัน firing rate จะเพิ่มขึ้นตามระดับของ tone จนถึงค่าสูงสุดค่าหนึ่ง เป็นจุดอิ่มตัว หมายความว่า ต่อให้ระดับของ tone จะเพิ่มมากกว่านี้ firing rate ก็ไม่เพิ่มไปมากกว่านี้ ดังรูป เราเรียกกราฟ rate-level function  นอกจากนี้ เส้นใยประสาทยังแสดงการเปลี่ยนแปลง firing rate ในโดเมนของเวลาอันเนื่องมาจากการเริ่มต้นเสียง หรือ onset ดังรูป 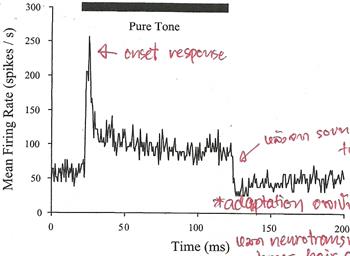 เห็นว่า ตอนเริ่มต้นเสียง firing rate มีค่าสูงสุด (onset response) และลดลงเมื่อเวลาผ่านไป และพอปิดเสียง firing rate จะตกลงต่ำว่า spontaneous rate ราว 100 ms Place Codingเนื่องจาก firing rate ของนิวรอนในเส้นประสาทการรับรู้เสียงถูกกำหนดโดยขนาดการสั่นของ basilar membrane ณ ตำแหน่งที่มันเชื่อมอยู่ ดังนั้น นิวรอนแต่ละตัวในเส้นประสาทมีความถี่ลักษณะเฉพาะประจำตัว และพวกมันก็อ่อนไหวต่อช่วงความถี่จำกัดรอบ ๆ ความถี่ลักษณะเฉพาะนี้เท่านั้น นอกจากนี้ การเพิ่มระดับเสียง ยังเพิ่ม firing rate ของนิวรอน จนกว่าจะถึงจุดอิ่มตัว ฉะนั้น วิธีหนึ่งที่ระบบการรับรู้เสียงจะใช้แทนหรือนำเสนอสเปกตรัมของเสียงคือในรูปของ firing rate ของนิวรอนที่แตกต่างกัน เช่น ถ้าเสียงใดมีองค์ประกอบความถี่ต่ำอยู่ นิวรอนที่มีความถี่ลักษณะเฉพาะใกล้เคียงกับองค์ประกอบตัวนั้นจะเพิ่ม firing rate เราเรียกการนำเสนอข้อมูลสเปกตรัมแบบนี้ว่า place code หรือ rate-place code เพราะข้อมูลสเปกตรัมถูกแทนด้วยรูปแบบของ firing rate ของกลุ่มนิวรอนPhase Locking & Temporal Codingplace coding มิได้เป็นเพียงวิธีการเดียวที่ characteristics ของเสียงจะถูกนำเสนอ การเปลี่ยนแปลงทางไฟฟ้าใน inner hair cell เกิดขึ้นเมื่อขนของมันเอียงชี้ไปทางด้านนอกของ cochlea เท่านั้น สมมติว่าตอนนี้ basilar membrane กำลังสั่นขึ้นลงตอบสนองต่อ pure tone ความถี่ต่ำ ขนหรือ stereocilia ของมันเอียงชี้สลับข้างไปมา ทีนี้ stereocilia จะเกิด depolarize ก็ต่อเมื่อมันเอียงไปทางด้านหนึ่งถูกมั้ยครับ นั่นเท่ากับ มันตอบสนองต่อบางเฟสเฉพาะของการสั่น หมายความว่า นิวรอนในเส้นประสาทการรับรู้เสียงจะผลิต spike ที่เฟสใดเฟสหนึ่งของ waveform สมบัตินี้เรียกว่า phase locking เพราะการตอบสนองของนิวรอนล็อกหรือจับคู่กับเฟสเฉพาะของการสั่น basilar membrane ดังรูป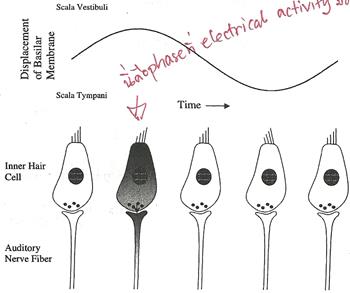 จากรูป สีเทา คือ เฟสที่ electrical activity มีค่าสูงสุด จากสมบัติ phase locking นี่เอง ได้บอกเป็นนัยถึงอีกวิธีหนึ่งที่ใช้นำเสนอหรือใช้แทนความถี่ในเส้นประสาทการรับรู้เสียง นั่นคือ การนำเสนอในรูปของ timing หรือ synchrony ของกิจกรรมในเส้นประสาท เช่น ถ้าเรากระตุ้นด้วย pure tone ความถี่ 100 Hz พบว่า นิวรอนมีแนวโน้มที่จะผลิต spike อยู่ห่างกันเป็นจำนวนเต็มเท่าของคาบของ pure tone กรณีนี้คือ 10 ms ทีนี้ firing rate ของนิวรอนไม่อาจมากกว่า 200 spike ต่อวินาที นี่จึงดูเหมือนเป็นขีดจำกัดของการใช้ประโยชน์จาก phase locking อยู่ที่ความถี่ประมาณ 200 Hz อย่างไรก็ตาม ถึงแม้ว่าเส้นใยแต่ละตัวจะไม่สามารถตอบสนองด้วยอัตราที่สูงพอต่อการเป็นตัวแทนทุกรอบของ waveform ที่เป็นตัวกระตุ้น แต่สามารถแก้ปัญหาได้ด้วยการเอาข้อมูลข่าวสารจากนิวรอนทุกตัวมารวมกันเพื่อใช้แทนความถี่ของ tone ความถี่สูง ดังรูป ความถี่ของ input เท่ากับ 250 Hz รูปกลางเป็นรูปแบบ spike ของนิวรอนตัวเดียว รูปล่างเป็นการรวม spike ที่ผลิตจากนิวรอน 500 ตัว  From Ear to Brainรูปต่อไปนี้แสดงเส้นทางส่งข้อมูลจากหูไปยังสมอง (ascending auditory pathways) เส้นประสาทนำข้อมูลจาก cochlea ส่งต่อไปยัง cochlea nucleus ซึ่งเป็นกลุ่มของนิวรอนใน brainstem (ก้านสมอง หรือ แกนสมอง) และส่งต่อไปยังนิวเคลียสต่าง ๆ ใน brainstem ดังรูป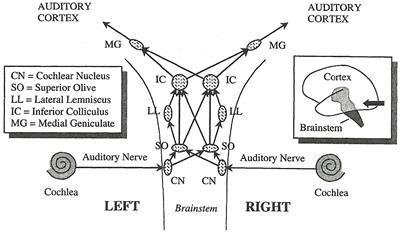 แต่ละช่วงของการส่งนั้น ข้อมูลเกี่ยวกับเสียงจะถูกประมวลผลโดยโครงข่ายนิวรอน เช่น SO ประมวลผลเกี่ยวกับการบอกตำแหน่งของแหล่งกำเนิดเสียง และ IC เกี่ยวกับการรับรู้ pitch ของเสียง เส้นใยประสาทจาก IC เชื่อมกับ (หรือสร้าง synapse กับ) MG ซึ่งเป็นส่วนหนึ่งของทาลามัส (thalamus) ในสมองส่วนกลาง โดยทาลามัสทำตัวประหนึ่งชุมสาย (relay station) สำหรับข้อมูลรับสัมผัส เส้นใยประสาทจาก MG ต่อกับ auditory cortex (เปลือกสมองทำหน้าที่ด้านการรับรู้เสียง) ซึ่งเป็นส่วนหนึ่งของ cerebral cortex (เปลือกสมองใหญ่) cerebral cortex คือ ส่วนที่มองเห็นเป็นรอยยับย่นนั่นแหละครับ จะปกคลุมพื้นผิวส่วนใหญ่ของสมอง ทำหน้าที่ข้องเกี่ยวกับการประมวลผลความคิดในระดับสูง รวมถึงฟังก์ชั่นรับสัมผัสกับการเคลื่อนไหวเบื้องต้น cortex นี้เป็นแผ่นนิวรอนบาง ๆ ความหนาเพียง 3 มิลลิเมตร มันขดพับจนดูเป็นรอยยับเพื่อจะได้มีพื้นที่ผิวมาก บางส่วนของ cerebral cortex รับข้อมูลจากระบบประสาทรับสัมผัส มีส่วนรับรูป รับเสียง เป็นต้น ส่วนรับเสียง หรือ auditory cortex อยู่บน temporal lobe (สมองกลีบขมับ) ซ่อนอยู่ที่รอยย่นใน cerebral cortex ที่เรียกว่า Sylvian fissure (ร่องซิลเวียน) ดังรูป 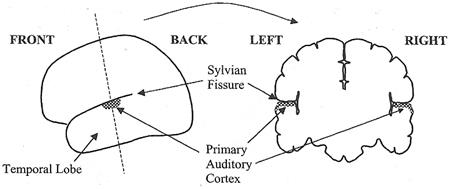 นอกจาก ascending auditory pathways ยังมี descending pathways ซึ่งข้อมูลถูกนำส่งจากศูนย์กลางประมวลผลเสียงระดับสูงในสมองลงไปยังระดับต่ำกว่า และลงไปถึงกระทั่ง cochlea และสามารถควบคุมการเคลื่อนที่ของ basilar membrane ดูเหมือนว่าระบบรับรู้เสียงถูกออกแบบมาเพื่อให้ศูนย์ประมวลมวลผลระดับสูงสามารถควบคุมกิจกรรมในระดับต่ำ และส่งผลกระทบต่อการประมวลผลเสียงได้ด้วยครับ
ขอบคุณมากครับ
โดย: hueristic (Hueristic
 ) วันที่: 4 พฤษภาคม 2556 เวลา:15:25:58 น. ) วันที่: 4 พฤษภาคม 2556 เวลา:15:25:58 น. |
บทความทั้งหมด
|
||||||||||||||||||||
 |


 ผู้ติดตามบล็อก : 85 คน [
ผู้ติดตามบล็อก : 85 คน [